Harakat potentsialining miqdoriy modellari - Quantitative models of the action potential
Yilda neyrofiziologiya, ning bir nechta matematik modellari harakat potentsiali ikkita asosiy turga bo'linadigan ishlab chiqilgan. Birinchi tur eksperimental ma'lumotlarni miqdoriy ravishda modellashtirishga, ya'ni tok va kuchlanish o'lchovlarini aniq ko'paytirishga intiladi. Taniqli Xojkin-Xaksli modeli aksonning Loligo kalmar bunday modellarga misol keltiradi.[1] Sifatida to'g'ri bo'lsa-da, H-H modeli har qanday qo'zg'aluvchan membranani aniq ta'riflab bermaydi, chunki u faqat ikkita ionni (natriy va kaliy) hisobga oladi, ularning har biri voltajga sezgir kanalga ega. Biroq, kabi boshqa ionlar kaltsiy muhim bo'lishi mumkin va barcha ionlar uchun juda xilma-xil kanallar mavjud.[2] Misol tariqasida yurak faoliyatining potentsiali kuchlanishga sezgir kaltsiy kanallari va har xil natriy / kaliy kanallari bo'lgan membranalarda har xil shakldagi ta'sir potentsiallari qanday hosil bo'lishi mumkinligini ko'rsatadi. Matematik modelning ikkinchi turi - bu birinchi turni soddalashtirish; Maqsad eksperimental ma'lumotlarni ko'paytirish emas, balki asabiy davrlarda harakat potentsialining rolini sifat jihatidan tushunishdir. Bunday maqsadda batafsil fiziologik modellar keraksiz darajada murakkablashishi va "daraxtlar uchun o'rmon" ni yashirishi mumkin. The FitzHugh-Nagumo modeli uchun o'rganiladigan ushbu sinfga xosdir qiziqish harakati.[3] O'qish odatda kuzatiladi tabiatda, masalan, ning sinxronlashtirilgan yoritilishida o't pashshalari, bu harakat potentsialining portlashi bilan muvofiqlashtirilgan;[4] individual neyronlarda tutilish ham kuzatilishi mumkin.[5] Kichkintoylarning xatti-harakatlarini tushunish uchun ikkala turdagi modellardan ham foydalanish mumkin biologik neyron tarmoqlari kabi markaziy naqsh generatorlari ba'zi bir avtomatik refleksli harakatlar uchun javobgardir.[6] Bunday tarmoqlar mushaklar qisqarishini muvofiqlashtirish uchun ishlatiladigan, masalan, yirtqichlardan qochish uchun nafas olish yoki tez suzish bilan shug'ullanadigan harakat potentsialining murakkab vaqtinchalik naqshini yaratishi mumkin.[7]
Xojkin-Xaksli modeli

1952 yilda Alan Lloyd Xodkin va Endryu Xaksli eksperimental kuchlanish-qisqich ma'lumotlarini aksonal membranaga moslashtirish uchun tenglamalar to'plamini ishlab chiqdi.[1][8] Model membrana sig'imini nazarda tutadi C doimiy; Shunday qilib, transmembran kuchlanishi V umumiy transmembran oqimi bilan o'zgaradi Mento'liq tenglamaga muvofiq

qayerda MenNa, MenKva MenL mahalliy natriy kanallari, kaliy kanallari va "oqish" kanallari (mos ravishda) orqali uzatiladigan oqimlar. Dastlabki muddat Menext kabi tashqi manbalardan keladigan oqimni ifodalaydi qo'zg'atuvchi postsinaptik potentsiallar dendritlardan yoki olim elektrodidan.
Model bundan keyin berilgan ion kanali to'liq ochiq yoki yopiq deb hisoblaydi; agar yopiq bo'lsa, uning o'tkazuvchanlik nolga teng, agar ochiq bo'lsa, uning o'tkazuvchanligi doimiy qiymatga ega g. Demak, ion kanali orqali aniq oqim ikki o'zgaruvchiga bog'liq: ehtimollik pochiq kanalning ochilishi va uning ionning muvozanat kuchlanishidan farqi, V − Vtenglama. Masalan, kaliy kanali orqali oqim quyidagicha yozilishi mumkin

ga teng bo'lgan Ohm qonuni. Ta'rifga ko'ra, aniq oqim oqimi yo'q (MenK = 0) transmembranali kuchlanish ushbu ionning muvozanat kuchlanishiga teng bo'lganda (qachon V = EK).
O'zlarining ma'lumotlariga aniq mos kelish uchun Xodkin va Xaksli ion kanallarining har bir turida bir nechta "eshiklar" bo'lgan deb taxmin qilishdi, shuning uchun kanal faqat barcha eshiklar ochiq va yopiq holda ochiq edi. Ular, shuningdek, eshikning ochilish ehtimoli boshqa eshiklarning ochiq bo'lishidan mustaqil deb taxmin qilishgan; bu taxmin keyinchalik inaktivatsiya eshigi uchun tasdiqlangan.[9] Xodkin va Xaksli voltajga sezgir kaliy kanalini to'rtta eshikli qilib modellashtirishdi; ruxsat berish pn bitta bitta eshikning ochilish ehtimolini bildiring, butun kanalning ochilish ehtimoli shunday to'rtta ehtimollikning hosilasi, ya'ni. pochiq, K = n4. Xuddi shu tarzda, voltajga sezgir bo'lgan natriy kanalining ehtimoli uchta shunga o'xshash ehtimollik eshiklari sifatida modellashtirilgan m va inaktivatsiya bilan bog'liq to'rtinchi eshik, ehtimollik h; shunday qilib, pochiq, Na = m3h. Har bir darvoza uchun ehtimolliklar bo'ysunadi deb taxmin qilinadi birinchi darajali kinetika

bu erda ham muvozanat qiymati mtenglama va bo'shashish vaqti doimiy constantm oniy kuchlanishga bog'liq V membrana bo'ylab. Agar V vaqt o'lchovida τ ga qaraganda sekinroq o'zgaradim, m ehtimollik har doim taxminan uning muvozanat qiymatiga teng bo'ladi mtenglama; ammo, agar V tezroq o'zgaradi, keyin m orqada qoladi mtenglama. Hodgkin va Xaksli o'zlarining kuchlanish-qisqich ma'lumotlarini o'rnatish orqali ushbu muvozanat qiymatlari va vaqt konstantalari harorat va transmembran kuchlanish bilan qanday o'zgarishini modellashtirishga muvaffaq bo'lishdi.[1] Formulalar murakkab va eksponent ravishda voltaj va haroratga bog'liq. Masalan, natriy kanalining faollashish ehtimoli uchun vaqt doimiysi h 3 sifatida o'zgaradi(− − 6.3) / 10 Selsiy harorati with va kuchlanish bilan V kabi

Xulosa qilib aytganda, Xojkin-Xaksli tenglamalari murakkab, chiziqli emas oddiy differentsial tenglamalar to'rtda mustaqil o'zgaruvchilar: transmembran kuchlanishi Vva ehtimolliklar m, h va n.[10] Ushbu tenglamalarning umumiy echimi topilmagan. Bunday noaniq dinamik tizimlarni o'rganish uchun kamroq ambitsiyali, ammo umuman qo'llaniladigan usul bu ularning xatti-harakatlarini sobit nuqta.[11] Ushbu tahlil Xojkin-Xaksli tizimi barqaror tinchlikdan -ga o'tishni ko'rsatmoqda yorilish tebranishlar rag'batlantiruvchi oqim sifatida Menext asta-sekin oshiriladi; diqqatga sazovor tomoni shundaki, akson stimulyator oqimi yanada oshgani sayin yana barqaror ravishda tinchlanadi.[12] Xodkin-Xaksli tenglamalari bashorat qilgan aksonlarning sifatli xatti-harakatlari turlarini yanada umumiy o'rganish amalga oshirildi.[10]
FitzHugh-Nagumo modeli
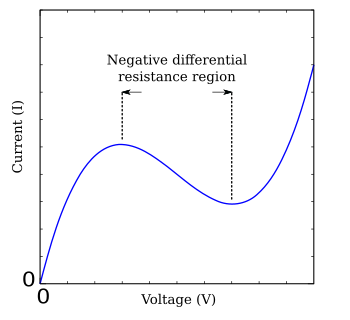
Xojkin-Xaksli tenglamalari murakkabligi sababli, sifat jihatidan o'xshash xulq-atvorni namoyish etadigan turli xil soddalashtirishlar ishlab chiqilgan.[3][13] The FitzHugh-Nagumo modeli bunday soddalashtirilgan tizimning odatiy namunasidir.[14][15] Asosida tunnel diodasi, FHN modeli faqat ikkita mustaqil o'zgaruvchiga ega, ammo to'liq Hodkin-Xaksli tenglamalariga o'xshash barqarorlikni namoyish etadi.[16] Tenglamalar


qayerda g (V) kuchlanishning funktsiyasi V o'rtada salbiy nishab mintaqasiga ega, uning yon tomoni bitta maksimal va bir minimal bilan chegaralangan (FHN-rasm). FitzHugh-Nagumo modelining juda ko'p o'rganilgan oddiy holati - bu tenglama bilan tavsiflangan Bonxeffer-van der Pol asab modeli.[17]


bu erda ε koeffitsienti kichik deb qabul qilinadi. Ushbu tenglamalarni ikkinchi darajali differentsial tenglamaga birlashtirish mumkin

Bu van der Pol tenglama chiziqli bo'lmagan matematikada ko'plab tadqiqotlarni rag'batlantirdi dinamik tizimlar. Op-amp harakat potentsialining FHN va van der Pol modellarini amalga oshiradigan sxemalar Keener tomonidan ishlab chiqilgan.[18]
Xodkin-Xaksli va FitzHyu-Nagumo modellarining duragaylari 1981 yilda Morris va Lekar tomonidan ishlab chiqilgan va muskul tolasi barnaklar.[19] Barnak fiziologiyasiga muvofiq, Morris-Lekar modeli Hodkin-Xaksli modelidagi voltajli natriy tokini kuchlanishga bog'liq kaltsiy oqimi bilan almashtiradi. Harakatsizlik yo'q (yo'q h o'zgaruvchan) va kaltsiy oqimi bir zumda muvozanatlashadi, shuning uchun yana vaqtga bog'liq bo'lgan ikkita o'zgaruvchi mavjud: transmembran kuchlanishi V va kaliy eshigi ehtimolligi n. Ushbu modelning yorilishi, mashq qilish va boshqa matematik xususiyatlari batafsil o'rganildi.[20]
Harakat potentsialining eng sodda modellari bu "yuvish va to'ldirish" modellari ("integratsiya va olov" modellari deb ham ataladi), ularda kirish signali chegaraga yetguncha ("to'ldirish" bosqichi) yig'ilib, puls va summani nolga qaytarish ("yuvish" bosqichi).[3][21][22] Ushbu modellarning barchasi namoyish etilishi mumkin qiziqish, bu odatda kuzatiladi asab tizimida.[3]
Hujayradan tashqari potentsiallar va oqimlar
Yuqoridagi modellar transmembrananing kuchlanishi va oqimini bitta membrana yamog'ida simulyatsiya qilgan bo'lsa, boshqa matematik modellar neyronni o'rab turgan ion eritmasidagi kuchlanish va oqimlarga taalluqlidir.[23] Bunday modellar hujayra ichidagi yozuvlarni amalga oshirishga imkon beradigan shisha pipet elektrod ixtiro qilinishidan oldin keng tarqalgan hujayradan tashqaridagi elektrodlardan ma'lumotlarni izohlashda yordam beradi.[24] Hujayradan tashqari muhit odatdagi izotrop sifatida modellashtirilishi mumkin ionli eritma; bunday echimlarda oqim quyidagilarga amal qiladi elektr maydon chiziqlari, ning doimiy shakliga ko'ra Ohm qonuni

qayerda j va E ni ifodalovchi vektorlardir joriy zichlik va elektr maydoni navbati bilan va qaerda σ bu o'tkazuvchanlik. Shunday qilib, j dan topish mumkin E, bu o'z navbatida yordamida topilishi mumkin Maksvell tenglamalari. Maksvell tenglamalarini nisbatan oddiy masalaga keltirish mumkin elektrostatik, ion kontsentratsiyasi juda sekin o'zgarganligi sababli (ga nisbatan yorug'lik tezligi ) uchun magnit ta'sir muhim bo'lishi. The elektr potentsiali φ (x) har qanday hujayradan tashqari nuqtada x yordamida hal qilish mumkin Yashilning o'ziga xosliklari[23]

bu erda integratsiya membrananing to'liq yuzasi ustida joylashgan; membranadagi holat, σichida va φichida membrana ichidagi o'tkazuvchanlik va potentsial, vatashqarida va φtashqarida mos keladigan qiymatlar membranadan tashqarida. Shunday qilib, membranadagi ushbu σ va φ qiymatlarni hisobga olgan holda hujayradan tashqari potentsial φ (x) har qanday pozitsiya uchun hisoblanishi mumkin x; o'z navbatida, elektr maydoni E va oqim zichligi j ushbu potentsial maydondan hisoblash mumkin.[25]

Shuningdek qarang
- Biologik neyron modellari
- GHK joriy tenglamasi
- Asabiy hisoblash modellari
- Tuzli o'tkazuvchanlik
- Bioelektronika
- Kabel nazariyasi
Adabiyotlar
- ^ a b v Xodkin AL, Xaksli AF, Katz B (1952). "Natriy va kaliy ionlari tomonidan Loligoning ulkan aksonining membranasi orqali o'tadigan oqimlar". Fiziologiya jurnali. 116 (4): 424–448. doi:10.1113 / jphysiol.1952.sp004717. PMC 1392213. PMID 14946713.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
Xodkin AL, Xaksli AF (1952). "Natriy va kaliy ionlari tomonidan Loligoning ulkan aksonining membranasi orqali o'tadigan oqimlar". Fiziologiya jurnali. 116 (4): 449–472. doi:10.1113 / jphysiol.1952.sp004717. PMC 1392213. PMID 14946713.
Xodkin AL, Xaksli AF (1952). "Loligo gigant aksonidagi membrana o'tkazuvchanligining tarkibiy qismlari". J Fiziol. 116 (4): 473–496. doi:10.1113 / jphysiol.1952.sp004718. PMC 1392209. PMID 14946714.
Xodkin AL, Xaksli AF (1952). "Loligo gigant aksonida membrana potentsialining natriy o'tkazuvchanligiga ikki tomonlama ta'siri". J Fiziol. 116 (4): 497–506. doi:10.1113 / jphysiol.1952.sp004719. PMC 1392212. PMID 14946715.
Xodkin AL, Xaksli AF (1952). "Membrana oqimining miqdoriy tavsifi va uning asab o'tkazuvchanligi va qo'zg'alishiga tatbiq etilishi". J Fiziol. 117 (4): 500–544. doi:10.1113 / jphysiol.1952.sp004764. PMC 1392413. PMID 12991237. - ^ Yamada WM, Koch C, Adams PR (1989). "Ko'p kanalli va kaltsiy dinamikasi". Yilda C. Koch, Men Segev (tahrir). Neyronal modellashtirish usullari: sinapslardan tarmoqlarga. Kembrij, Massachusets: Bradford Book, The MIT Press. 97-133 betlar. ISBN 978-0-262-11133-1.CS1 maint: bir nechta ism: mualliflar ro'yxati (havola)
- ^ a b v d Hoppensteadt FK (1986). Neyronlar matematikasiga kirish. Kembrij: Kembrij universiteti matbuoti. ISBN 978-0-521-31574-6.
- ^ Xanson, F.E .; Case, J.F .; Bak, E .; Buck, J. (1971). "Yangi Gvineya Firefly-da sinxronizatsiya va flesh o'yinlar". Ilm-fan. 174 (4005): 161–164. Bibcode:1971Sci ... 174..161H. doi:10.1126 / science.174.4005.161. PMID 17742039.
- ^ Guttman R, Feldman L, Jacobsson E (1980). "Kalamar aksonining chastotasini o'rganish". J. Membr. Biol. 56 (1): 9–18. doi:10.1007 / BF01869347. PMID 7441721.
- ^ PA olish (1989). "Kichik asab tarmoqlarini qayta qurish". Yilda C Koch va men Segev (tahr.) Neyronal modellashtirish usullari: sinapslardan tarmoqlarga. Kembrij, Massachusets: Bradford Book, The MIT Press. 171-194 betlar. ISBN 978-0-262-11133-1.
- ^ Hooper, Scott L. "Markaziy naqsh ishlab chiqaruvchilari". Embrion ELS (1999) http://www.els.net/elsonline/figpage/I0000206.html[doimiy o'lik havola ] (2 ning 2) [2006.06.06 11:42:28 AM] Onlayn: 2007 yil 27-noyabrda kirish [1].
- ^ Nelson ME, Rinzel J (1994). "Xojkin-Xaksli modeli" (PDF). Bower J, Beeman D (tahrir.). GENESIS Kitobi: GEneral NEural SImulation System yordamida real neyron modellarni o'rganish. Nyu-York: Springer Verlag. 29-49 betlar.
- ^ Armstrong CM, Bezanilla F, Rojas E (1973). "Pronaz bilan to'ldirilgan kalamar aksonlarida natriy o'tkazuvchanlik inaktivatsiyasini yo'q qilish". J. Gen. Fiziol. 62 (4): 375–391. doi:10.1085 / jgp.62.4.375. PMC 2226121. PMID 4755846.
Rojas E, Rudi B (1976). "Natriy o'tkazuvchanligini inaktivatsiyasini parchalangan asab tolalarida o'ziga xos proteaz bilan yo'q qilish Loligo". J. Fiziol. 262 (2): 501–531. doi:10.1113 / jphysiol.1976.sp011608. PMC 1307656. PMID 994046. - ^ a b Sato S, Fukai H, Nomura T, Doi S (2005). "Xojkin-Xaksli tenglamalarining bifurkatsion tahlili". Reeke GN, Poznanski RR, Lindsay KA, Rozenberg JR, Sporns O (tahr.). Nörobilimlerde modellashtirish: Biologik tizimlardan neyromimetik robototexnika (2-nashr). Boka Raton: CRC Press. 459-478 betlar. ISBN 978-0-415-32868-5.
- ^ Gukkenxaymer J, Xolms P (1986). Vektorli maydonlarning chiziqli bo'lmagan tebranishlari, dinamik tizimlari va bifurkatsiyalari (2-nashr, qayta ko'rib chiqilgan va tuzatilgan.). Nyu-York: Springer Verlag. 12-16 betlar. ISBN 978-0-387-90819-9.
- ^ Sabah NH, Spangler RA (1970). "Hodgkin-Xaksli modelining kalamar giganti akson uchun takroriy javobi". Nazariy biologiya jurnali. 29 (2): 155–171. doi:10.1016/0022-5193(70)90017-2. PMID 5500466.
Evans JW (1972). "Nerv akson tenglamalari. I. Chiziqli yaqinlashishlar". Indiana Univ. Matematika. J. 21 (9): 877–885. doi:10.1512 / iumj.1972.21.21071.
Evans JW, Feroe J (1977). "Nerv impulsining mahalliy barqarorlik nazariyasi". Matematika. Biosci. 37 (1–2): 23–50. doi:10.1016/0025-5564(77)90076-1. - ^ Fitz Xyu R (1960). "Hodkin-Xaksli asab tenglamalarida ostonalar va platolar". J. Gen. Fiziol. 43 (5): 867–896. doi:10.1085 / jgp.43.5.867. PMC 2195039. PMID 13823315.
Kepler TB, Abbott LF, Marder E (1992). "Supero'tkazuvchilarga asoslangan neyron modellarini qisqartirish". Biologik kibernetika. 66 (5): 381–387. doi:10.1007 / BF00197717. PMID 1562643. - ^ Fitz Xyu R (1961). "Nerv membranasining nazariy modellaridagi impulslar va fiziologik holatlar". Biofizika jurnali. 1 (6): 445–466. Bibcode:1961BpJ ..... 1..445F. doi:10.1016 / S0006-3495 (61) 86902-6. PMC 1366333. PMID 19431309.
- ^ Nagumo J, Arimoto S, Yoshizava S (1962). "Nerv aksonini simulyatsiya qiluvchi faol impuls uzatish liniyasi". IRE ishi. 50 (10): 2061–2070. doi:10.1109 / JRPROC.1962.288235.
- ^ Fitz Xyu R (1969). "Axtsitatsiya va asabda tarqalishning matematik modellari". HP Shvann (tahr.) Da. Biologik muhandislik. Nyu-York: McGraw-Hill. 1-85 betlar.
- ^ Bonxeffer KF (1948). "PASSIV DEMIRNI NERVNI QO'LLASH UChUN O'RNAK sifatida faollashtirish". J. Gen. Fiziol. 32 (1): 69–91. doi:10.1085 / jgp.32.1.69. PMC 2213747. PMID 18885679.
Bonxeffer KF (1953). "Modelle der Nervenerregung". Naturwissenschaften. 40 (11): 301–311. Bibcode:1953NW ..... 40..301B. doi:10.1007 / BF00632438.
van der Pol B (1926). "Bo'shashish-tebranishlar to'g'risida". Falsafiy jurnal. 2: 978–992.
van der Pol B, van der Mark J (1928). "Yurak urishi gevşeme tebranishi va yurakning elektr modeli sifatida qabul qilinadi". Falsafiy jurnal. 6: 763–775.
van der Pol B, van der Mark J (1929). "Yurak urishi gevşeme tebranishi va yurakning elektr modeli sifatida qabul qilinadi". Arch. Nerl. Fiziol. 14: 418–443. - ^ Keener JP (1983). "Van der Pol va Fitsz-Nagumo tenglamalari uchun analog sxema". IEEE tizimlari, inson va kibernetika bo'yicha operatsiyalar. 13 (5): 1010–1014. doi:10.1109 / TSMC.1983.6313098.
- ^ Morris C, Lecar H (1981). "Barnacle gigant mushak tolasidagi kuchlanish tebranishlari". Biofizika jurnali. 35 (1): 193–213. Bibcode:1981BpJ .... 35..193M. doi:10.1016 / S0006-3495 (81) 84782-0. PMC 1327511. PMID 7260316.
- ^ Rinzel J, Ermentrout GB (1989). "Asab qo'zg'aluvchanligi va tebranishlarini tahlil qilish". Yilda C. Koch, Men Segev (tahrir). Neyronal modellashtirish usullari: sinapslardan tarmoqlarga. Kembrij, Massachusets: Bradford Book, The MIT Press. 135–169 betlar. ISBN 978-0-262-11133-1.
- ^ Keener JP, Hoppensteadt FC, Rinzel J (1981). "Nerv membranasini tebranuvchi kirishga javob berishning birlashtirilgan va olovli modellari". Amaliy matematika bo'yicha SIAM jurnali. 41 (3): 503–517. doi:10.1137/0141042.
- ^ Sejnar, Pavel; Vishata, Oldxich; Kukal, Jaromir; Beranek, Martin; Valish, Martin; Procházka, Aleš (2020). "Barcha qismlari biologik jihatdan izohlangan qo'zg'atuvchi va inhibitor neyronlarning oddiy kondensator-kommutatorli modeli kirish rejimiga bog'liq bo'lgan xaotik tebranishlarga imkon beradi". Ilmiy ma'ruzalar. 10 (1): 7353. doi:10.1038 / s41598-020-63834-7. PMID 32355185.
- ^ a b Stivens, Charlz F. (1966). Neyrofiziologiya: boshlang'ich. Nyu-York: Jon Vili va o'g'illari. pp.161 –173. LCCN 66015872.
- ^ Ling G, Jerar RW (1949). "Baqa sartorius tolalarining normal membrana potentsiali". J. hujayra. Komp. Fiziol. 34 (3): 383–396. doi:10.1002 / jcp.1030340304. PMID 15410483.
- ^ Lorente de No R (1947). "Asablar fiziologiyasini o'rganish". Stud. Rokfeller Inst. Med. Tadqiqot. 132: Chap. 16.
Mauro A (1960). "Hajmi o'tkazgichlarda elektrofiziologik potentsialga taalluqli nozik generatorlarning xususiyatlari". J. neyrofiziol. 23 (2): 132–143. doi:10.1152 / jn.1960.23.2.132.
Woodbury JW (1965). "3-bob: Ovoz o'tkazgichidagi potentsial". TC Ruchda; HD Patton (tahrir). Fiziologiya va biofizika. Filadelfiya: W. B. Saunders Co.
Qo'shimcha o'qish
- Glass L, Mackey MC (1988). Soatlardan tartibsizlikka: hayot ritmlari. Prinston, Nyu-Jersi: Prinston universiteti. ISBN 978-0-691-08496-1.